
Título
Autor
Fecha
Lugar de Realización
Texto
Premio AANC 2002
El Espacio Perforado Anterior y Sus Zonas Aledañas. Con Consideraciones Funcionales Parte II
Horacio Fontana, Héctor Belziti y Flavio Requejo
Servicio de Neurocirugía del Hospital Municipal de San Isidro.
Correspondencia: Arrotea 756, Depto A (1406) Buenos Aires
RESUMEN
Los vasos que perforan el EPA provienen de la carótida, cerebral anterior y medía y coroidea anterior. Tienen zonas de penetración típicas pero parcialmente superpuestas y su distribución obedece a razones topográficas generadas por la estructura del EPA y condicionadas filogenéticamente.
Aunque no correcta, la denominación lenticuloestriada para los ramos silvianos, ha sido impuesta por el uso.
Funcionalmente, se distinguen tres sistemas no separables anatómicamente, el pálido ventral, el núcleo basal y la amígdala-séptum, que confieren al EPA su actividad relevante en procesos de memoria, atención, conducta y personalidad.
Palabras clave: núcleo basal, sustancia innominada, vasos perforantes anteriores.
ABSTRACT
The EPA's perforating vessels are originated from the carotid, anterior cerebral and middle cerebral arteries. They have tipícal entry zones that overlap partially and their distribution depends on philogenetically conditioned topographical relations. Though incorrect, the naming lenticulo striate for the perforating branches from the silvian arter y, has been imposed by its use.
'There are three functional systems at the EPA„ that cannot be anatomically ditinguished: the ventral pallidum, the basal nucleus and the amygdala- septum, that give the EPA its relevance in processes of memory, attention, behavior and personality.
Key words: anterior perforating arteries, basal nucleus, substancia innominata.
VASOS PERFORANTES
El EPA es el hilio de las estructuras profundas del telencéfalo.
La porción terminal de la carótida interna y las iniciales de la cerebral anterior y media circulan por debajo de su superficie, conformando una especie de letra T cuyo tramo horizontal está ubicado algo oblicuamente hacia atrás y afuera. Ramos de estos tres vasos y de la coroidea anterior, penetran a través de su sustancia.
Analicemos las áreas de penetración de los ramos de los distintos vasos implicados.
En un preparado del espacio perforado anterior con sus vasos (Fig. 1), vemos la carótida, cerebral anterior, ambas A2 y una gruesa comunicante anterior. La arteria recurrente de Heubner, naciendo en la zona de la arteria comunicante anterior.
Podemos observar que la arteria comunicante posterior no emite ramos para el espacio perforado anterior, ya que todas penetran por dentro de la cintilla óptica, barrera natural entre dos sistemas (diencefálico y telencefálico).

Fig. 1. Vista basal del cerebro a nivel del EPA izquierdo. El nervio óptico izquierdo ha sido reclinado levemente hacia abajo y la terminación de la carótida (flecha celeste) hacia fuera. te: polo temporal izq. parcialmente resecado; to: trígono olfatorio; mm: tubérculos mamilares. Flechas verde claro: cerebral anterior. PorciónAl marcada con una flecha; porciones A2 una flecha cada una, unidas por una comunicante anterior gruesa (*). Flechas azules: arteria recurrente de Heubner que nace en el origen de A2 y penetra dentro del trígono olfatorio, próximo a la cisura entorrinal. Flecha marrón: cintillo óptica izquierda. Flecha negra: arteria comunicante posterior gruesa, de la que se ven nacer ramos perforantes de los cuales ninguno va al EPA, ya que entran por dentro de la cintilla óptica, que es una barrera natural entre dos sistemas: telencefálico porfuera, diencefálico por dentro.
Reclinando carótida y comunicante posterior en el mismo espécimen hacia adentro (Fig. 2), vemos de nuevo la cintilla óptica y los ramos perforantes que penetran ahora por fuera de ella, en el EPA, que provienen de la arteria coroidea anterior, que cruza muy oblicuamente la cintilla.
También vemos ramos que nacen de la silviana que penetran en pleno espacio, o en la cisura entorrinal y ramos de la bifurcación carotídea.
En la figura 3 podemos ver ramos silvianos que penetran en la parte más lateral del espacio perforado y otro que en cambio se dirige hacia adentro para penetrar en el área de entrada de la recurrente de Heubner, justo por detrás del trígono olfatorio. También se observan múltiples ramos delgados de Al que van a penetrar por delante y dentro de la cintilla y de la bandeleta de Broca. De la arteria comunicante anterior se ve salir un solo ramo que, como corresponde, no va al espacio perforado anterior.
En otro espécimen (Fig. 4), vemos la arteria recurrente y los ramos cortos de Al; ramos carotídeos nacidos inmediatamente antes de la bifurcación; la coroidea, con ramos que penetran en la parte posterior del espacio y de la cisura entorrinal y ramos silvianos al espacio mismo y a la cisura entorrinal.

Fig. 2. Mismo espécimen que el anterior, rotado casi 90 grados. La carótida terminal ha sido reclinada hacia adentro (flecha celeste). Flecha celeste corta: ramos perforantes que nacen de la carótida terminal. Flechas amarillas• larga: arteria coroidea anterior; cortas: sus ramos perforantes para la zona posterior del EPA. Flecha verde claro: Cintilla óptica. La arteria coroidea anterior la va a cruzar casi longitudinalmente. Los ramos coroideos perforantes por fuera de la cintilla, van al EPA. Los que da luego de cruzarla, van al tálamo y c. geniculado externo. Se pueden ver también, las ramas perforantes de la comunicante posterior que, como dijimos, no van al EPA. Flechas lila: arteria cerebral media y sus ramos perforantes. La flecha más corta, señala ramos que penetran en la porción temporal de la cisura entorrinal. T: túber; nervio óptico izquierdo; m: tubérculo mamilar.

Fig. 3. Mismo espécimen que el anterior. La carótida interna y su bifurcación han sido desplazados hacia atrás, para poder ver la parte anterior del EPA. Flechas azules: larga: arteria recurrente de Heubner, penetrando en la cisura entorrinal levemente por dentro del trígono olfatorio; cortas: ramos cortos de Al que penetran levemente por delante de la cintilla y quiasma. Flechas lila: ramos perforantes de la arteria cerebral media (lentículo estriadas en forma genérica). Una se dirige a la zona de entrada de la arteria recurrente. El racimo lateral, penetra en la porción más lateral de la cisura entorrinal. Obsérvese que pequeños ramos no señalados, penetran en pleno espacio perforado.

Fig. 4. Otro hemisferio izquierdo. La carótida y sus ramos han sido reclinados hacia delante. Flecha celeste: perforantes de la porción terminal de la carótida interna. Penetran en el centro del EPA. Flechas amarillas: arteria coroidea anterior y sus ramos perforantes entrando en la parte posterior del EPA. Obsérvese la relación de la coroidea anterior con la cintilla óptica y cómo la cruza. Flechas azules: se observan ramos cortos de Al , penetrando por delante de la cintilla y quiasma, seguramente también por delante de la bandeleta de Broca. La arteria de Heubner está parcialmente oculta por el trayecto de A1
Abbie1 (Fig. 5) atribuye una zona de penetración más bien lateral a los ramos de la arteria de Heubner, pero nosotros en los casos presentados, la hemos visto entrar más medialmente, por detrás del trígono olfatorio, así que su zona de entrada es algo más amplia, como también lo ha hecho notar el grupo de Rothon2. La zona de penetración silviana se superpone en parte a ella y tiene vasos que perforan en el espacio, en la cisura entorrinal, o por fuera de ella, en el lóbulo piriforme, y tienen territorios diferentes. Los ramos coroideos entran por detrás en el espacio y la cisura, Salvo el grupo de Rothon, casi ningún autor hace referencia a los ramos carotídeos, que sin embargo son importar!-tes quirúrgicamente, por su posible daño al clipar aneurismas de la bifurcación. Estos vasos pueden abastecer territorios atribuidos en la literatura previa a las arterias cerebral anterior, media o coroidea anterior.
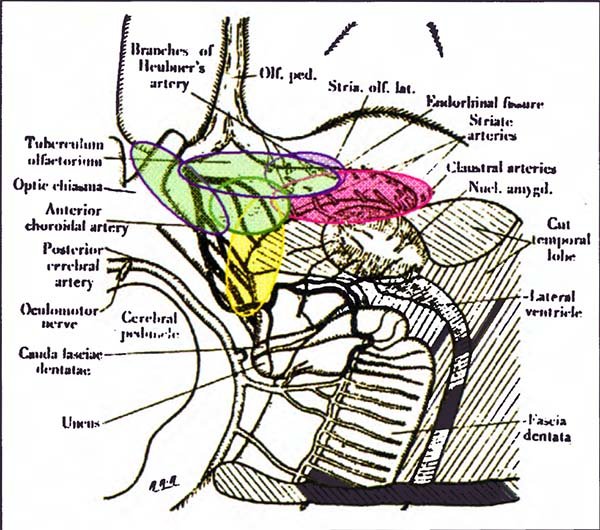
Fig. 5. Esquema modificado de la penetración de los distintos vasos perforantes, según Abbiel (1934), Óvalos azules: zonas de penetración de los ramos de la cerebral anterior, El óvalo más pequeño, encierra los ramos de la arteria de Heubner según Abbie, Nosotros hemos visto que esta arteria puede penetrar más adentro, siempre en la vecindad de la porción anterior de la cisura entorrinal (óvalo anterior, más grande), Coincidimos en esto, con el grupo de Rothon2. El óvalo oblicuo, paralelo a la cintilla óptica, es la zona de penetración de los ramos cortos de A1, como vimos, Óvalo rosa: zona de penetración de las lentículo estriadas, ramas de la cerebral media. En el dibujo parecen penetrar todas en la cisura entorrinal, pero hemos visto que también pueden penetrar en el centro del espacio. Para Abbie, los vasos que penetran por fuera de la cisura, no van al cuerpo estriado, sino al claustro y las otras cápsulas, Óvalo celeste: zona de penetración de los ramos de la carótida terminal. Son los menos constantes2 y su territorio puede ser suplido por cualquiera de los otros, Óvalo amarillo: territorio de penetración de los ramos coroideos.
Los ramos finos de Al penetran siempre por delante de la bandeleta de Broca, según vimos.
Para Abbie, los vasos que penetran en pleno espacio perforado, cualquiera sea su origen, irrigan las partes antiguas del estriado, es decir el pálido y la cápsula en sus porciones más bajas, Las que penetran en su periferia, es decir, en la vecindad de la cisura entorrinal, irrigan las estructuras nuevas, caudado, putamen y porciones altas de la cápsula interna y las que lo hacen por fuera de ella, abastecen el claustro y las otras cápsulas, Veamos nuestro esquema inicial (Fig. 6) para comprender desde el punto de vista topográfico, esta notable observación de Abbie. Las porciones anteriores y laterales de la cisura entorrinal se ubican exactamente por debajo de las porciones nuevas del estríado. Las porciones más posteriores del mismo, tendrán que ser irrigadas por los vasos más laterales (ver más abajo) que seguirán un curso hacia atrás y arriba dentro del parénquima.

Fig. 6. Esquema inicial, mostrando el espacio perforado y la relación de la cisura entorrinal con las estructuras profundas. En líneas de puntos azul se ve la zona que corresponde a la cisura entorrinal. Por transparencia se ve que el estriado está ubicado encima de sus porciones anteromedial, anterolateral y parte de la posterolateral, lo que hace muy verosímil la teoría de Abbie (ver texto). El vértice anterior corresponde a la zona de penetración de la arteria recurrente y en profundidad, con la cabeza del caudado y el brazo anterior de la cápsula interna, El vértice lateral corresponde a la penetración de las lentículo estriadas laterales que van al putamen, cápsula interna alta y cuerpo del caudado, El centro del EPA corresponde al pálido y parte baja de la cápsula interna, como así también al asa peduncular.
La superposición de los territorios está indicando que puede haber predominio de un vaso sobre otro en determinado paciente, lo que se expresa por la constancia de territorios pero con cierta variabilidad de orígenes, ya observada por Duret3.
En la figura 7 vemos un ejemplo. La coroidea anterior emite varios vasos que se dirigen a la parte anterior del espacio, además de sus ramos habituales posteriores por fuera de la cintilla. Podemos apreciar también la vena silviana profunda con venas lenticuloestriadas aferentes y su continuación en la vena basal de Rosenthal.

Fig. 7. Hemisferio izquierdo. Ejemplo de suplencia de un territorio por otro vaso. Flecha celeste: carótida terminal. Flecha lila: arteria cerebral media. Flecha negra: arteria comunicantecerebral posterior. Flechas amarillas: arteria coroidea y sus ramas. Obsérvese cómo hay ramos que se dirigen hacia delante para suplir el territorio de la carótida o de la cerebral media, además de los característicos ramos para la porción posterior del EPA. Flechas naranja: vena silviana profunda recibiendo venas lenticuloestriadas, vasos que también "perforan" el EPA, aunque en sentido inverso. Se ve la continuación con la vena basal de Rosenthal al cruzar la cintilla óptica. Flecha jaspeada: cintilla óptica.

Fig. 8. Trayecto intraparenquimatoso de los vasos lenticuloes triados en un hemisferio izquierdo (inyección con gelatina). Se puede ver que los ramos más internos son los más anteriores. Las flechas azules muestran una típica curva hacia atrás que hace el grupo y que atribuimos inicialmente, nos parece que en forma errónea, al pasaje de la comisura anterior (flecha roja). Laflecha lila de la izquierda muestra algunos vasos saliendo de un tronco común, lo que no es raro. La flecha lila de la derecha muestra un ramo lateral que hace un trayecto francamente recurrente antes de entrar al EPA.
El trayecto profundo de los vasos lentículo estriados es bastante característico y constante (Fig. 8). Los más anteriores son los más internos y más cortos. Además, hacen una curva hacia atrás, que interpretamos inicialmente como causada por el paso de la comisura anterior que cruza en esta zona, pero luego vimos que tanto los vasos que pasan por delante de ella, como los que lo hacen por detrás, sufren la misma desviación, así que debería atribuirse a otras razones, quizá el pasaje del fascículo unciforme.
Se ve también cómo pueden nacer varias perforantes de un tronco común, una disposición frecuente2 y el largo trayecto recurrente de las más laterales.
En la figura 9 vemos un esquema redibujado de Gillilan4 y modificado para permitir la presentación de los vasos por planos.
La nomenclatura de los ramos perforantes es variada y algo contradictoria. Resumimos los aportes de los distintos autores en el cuadro 1.
En 1874, Duret3 describió para la cerebral anterior ramas cortas y una o dos arterias retrógradas. A esta arteria recurrente se refiere ese mismo año Heubner, quedando su nombre ligado a la misma. Las ramas de la cerebral media, las dividió Duret muy lógicamente en internas o lenticulares y externas o lenticuloestriadas.

Fig. 9. Esquema del trayecto de los ramos perforantes por planos progresivos, redibujado y modificado de L. Gilligan4, (1968).
A. Car carótida interna; Ca: cerebral anterior; CM: cerebral media; Pu: putamen; ge: cuerpo geniculado externo.
B. Ramos de la coroidea. Al EPA entran ramos para el pálido, putamen y cápsula interna. Fuera de EPA, de vasos para el cuerpo geniculado, tálamo y plexo coroideo.
C. Ramos silvianos (lentículoestriadas).
D. A recurrente de Heubner.
Cuadro 1. Nomenclatura de los ramos perforantes

En 1910, Mouchet5 habla de estriadas anteriores y posteriores
En 1932, Abbie1 habla de arteria de Heubner y divide las arterias estriadas en tres grupos, de acuerdo a la zona que perforan, como hemos visto. Andersen6, en la primera descripción radiológica precisa de los perforantes silvianos, habla de lenticuloestriadas mediales y laterales, hito histórico, que marcó la denominación de estos vasos.
Lazorthes7 en 1961, habla de centrales cortas y central larga para los ramos de Al, y llama, siguiendo a Foix y Levy, palidales externas a los perforantes internos y putámino cápsulo caudadas a los externos provenientes de la silviana.
En 1968, Westberg8, denomina lenticuloestriadas mediales a la arteria de Heubner y lentículoestriadas laterales a los ramos silvianos, generando buena parte de la confusión reinante.
Gillilan4 en 1968, denomina estriadas media-les a la arteria de Heubner y estriadas laterales a los ramos silvianos, reconociendo en éstos, un grupo medial y otro lateral.
En 1976 Schlessinger9, habla de estriocapsulares mediales y laterales distinguiendo en éstas un grupo pre y otro postcomisural.
Salamon5 en 1978, sigue la descripción de Duret, pero denomina lenticuloestriadas a todo el grupo de la cerebral media.
En 1984, el grupo de Rothon2 habla de ramas de Al y arteria de Heubner para la cerebral anterior y denomina lenticuloestriadas a las de la cerebral media dividiéndolas en tres grupos de acuerdo al sector de emergencia desde el vaso madre.
Vemos que hay una coincidencia importante en los conceptos, pero un uso errático de los términos medial y lateral y lenticuloestriada. Este término, restringido en su acepción inicial, por Duret, se ha extendido por el uso neurorradiológico a todos los perforantes silvianos y está impuesto por el uso también en la Neurocirugía.
El límite externo del espacio perforado coincide con el limen de la ínsula, que representa una elevación correspondiente al fascículo uncinado (Fig. 10 A y B).
En la figura 10 C se puede observar la disección del fascículo unciforme junto con el longitudinal inferior. Este pliegue genera el punto más estrecho de la porción horizontal de la cisura de Silvio que, luego de trasponerlo, va a volver a ampliarse en la cisterna de la ínsula.

Fig. 10. Fascículo unciforme y pliegue falciforme.
A. El fascículo unciforme y frontotemporal une la región frontoorbitaria y porciones de F2 al polo temporal y produce la elevación de la corteza llamada pliegue falciforme, que se ve afuera, como el umbral de la ínsula (flecha roja). (Re-dibujado e E. Crosby10).
B. Umbral de la ínsula.
C. Disección de fascículo unciforme (flecha roja) y fascículo frontooccipital inferior (flecha verde) en un hemisferio izquierdo. Esta zona es el punto más estrecho (S) de la cisterna silviana, que se ensancha como un reloj de arena por dentro (porción horizontal correspondiente al EPA) y por fuera, en la cisterna de la ínsula.

Fíg. 11. Angiografía derecha de frente donde se visualizan vasos y elementos del EPA. Asteriscos rojos: puntos de entrada de los vasos lentículo estriados en el EPA. Estos vasos realizan un trayecto cisternal desde la arteria silviana, variablemente recurrente, más pronunciado cuanto más laterales. En su trayecto parenquimatoso, un curso curvo de convexidad externa, paralelo a las láminas medulares del nervio lenticular. Flechas blancas: señalan el trayecto de la arteria recurrente de Heubner. Flechas amarillas: larga, trayecto de la arteria coroidea anterior; cortas, vasos perforantes ramos de esta arteria. Forma roja: corte del fascículo unciforme ubicado entre las lentículoestriadas más laterales y los primeros vasos insulares. Línea negra: estimación de la superficie cerebral en el EPA y pliegue falciforme.
Aunque se trata de una estructura nerviosa, ha sido traída a la discusión en este tramo de la presentación, porque su posición puede ser calculada en una angiografía de frente (Fig. 11), en la que vemos la inflexión de las lenticuloestriadas en el momento que penetran en el espacio perforado. Son vasos casi siempre recurrentes en su trayecto extraparenquimatoso, más marcadamente cuanto más laterales. Vemos la posición del fascículo unciforme, y cómo es delimitado entre el comienzo de los vasos insulares de la silviana y la lentíiculoestriadas más externas. Se visualiza también la recurrencial de Heubner; la coroidea anterior con ramos geniculados y la superficie del cerebro en el pliegue falciforme y el espacio perforado.
FILOGENIA
En la figura 12, tomada de Abbie1, vemos esquemáticamente representada la evolución filo-genética del EPA, que es una de las estructuras más antiguas del telencéfalo, y por lo tanto, presente en todos los vertebrados.

Fig. 12. Evolución filogenética del EPA. Redibujado y levemente modificado de Abbie, 1932. Flechas verdes: cisura entorrinal; to: tubérculo olfatorio.
A. Cocodrilo. Dos vasos homólogos de cerebral anterior y media, uno lateral y otro medial forman una red sobre el tubérculo olfatorio.
B. Oveja. El crecimiento lateral del neocórtex comienza a deformar transversalmente el EPA y produce el desarrollo de un vaso neocortical lateral marcado con la flecha rosa (cerebral media).
C. Primate. Las dos arterias neocorticales están desarrolladas. El EPA ha adquirido su disposición predominantemente transversal. De la red inicial se diferencia la recurrente de Heubner predominando sobre la cerebral media. Las flechas rosa muestran vasos estriados silvianos entrando en la parte posterior de la cisura entorrinal.
En un reptil, la zona que estudiamos constituye casi todo el telencéfalo y está irrigada por un vaso medial y otro lateral que representarían las arterias cerebral anterior y media primitivas. Se destacan el tubérculo olfatorio y la cisura entorrinal.
En una oveja, el vaso lateral tiene un gran requerimiento de la neocorteza, a la vez que el espacio perforado comienza a plegarse, como lo muestra la angulación de la cisura entorrinal. La red que irriga esta zona, ha sufrido pocos cambios. Proporcionalmente la zona es más pequeña en el primate, lo mismo que el tubérculo olfatorio, y se ha plegado completamente en forma transversal, por el gran desarrollo del neocórtex, que provoca un desarrollo semejante de las arterias cerebral media y anterior.
En el espacio perforado, alguna de las ramas transversales que lo irrigan se hipertrofian como arteria de Heubner y usurpan parcialmente el territorio perforante de la cerebral media, que también emite ramos para la cisura entorrinal.
CONSIDERACIONES FUNCIONALES
Desde el punto de vista funcional encontramos en el espacio perforado anterior, tres sistemas imposibles de separar anatómicamente, lo que ha hecho muy dificil su estudio: el núcleo basal de Meynert asociado a la sustancia innominada, el continuo amígdala septum y el pálido ventral11.
Cuadro 2. Los sistemas funcionales del espacio perforado anterior

La sustancia innominada forma islotes de sustancia gris unidos por pequeñas hileras celulares que rellenan todo este espacio fibroso que venimos estudiando12,13. Sus límites son irregulares y aproximados (Fig. 13). Por arriba el núcleo lenticular. Por debajo, la superficie del espacio perforado y por dentro se continúa insensiblemente con los núcleos del túber. Se le reconocen dos prolongaciones: una que sigue el lecho de la estría terminal y otra que se extiende por la lámina medular externa del núcleo lenticular para alcanzar a veces la cápsula interna12,13. Marcamos en este corte el sistema de fibras longitudinales que circulan por el hipotálamo lateral, límite natural entre las dos regiones.
En la figura 14, que compara nuestra disección con un corte horizontal de Foix y Nicolesco, resumimos las nociones estructurales discutidas hasta ahora y las relacionamos a la sustancia innominada. Es notable y bella la coincidencia entre las estructuras marcadas en ambas imágenes.
En un esquema levemente modificado de Haber" (Fig. 15), vemos el estriado ventral, correspondiente al "fundus striati" de Hassler14, que corresponde a las porciones basales de putamen y caudado y al núcleo accumbens. El pálido ventral ocupa la parte inferomedial del pálido externo y se extiende por debajo de la comisura anterior.
En el cuadro 3, observamos las conexiones del pálido ventral, que serían una vía de descarga hacia centros inferiores (¿respuestas motoras?) de las estructuras límbicas, y menos probablemente, según investigaciones recientes, de modulación cortical prefrontal desde el sistema límbico (flecha punteada)15. La vía descendente pasa en parte por el asa lenticular.
En el cuadro 4 se observa la constitución del ganglio basal de Meynert y los tres núcleos de que está constituido según Arendt16, cuya ubicación es mostrada en esquemas en la figura 16.

Fig. 13. Extensión de la sustancia innominada marcada en un corte coronal que pasa por el agujero de Monro (tomado de Foix y Nicolesco). El EPA es la superficie de una placa de sustancia gris surcada por los múltiples fascículos que hemos mostrado, de 1 a 1,5 cm de espesor, extendida por debajo del núcleo lenticular y cápsula interna. Esta placa de sustancia gris recibe el nombre de sustancia innominada. Está constituida por grupos pequeños de células grandes y medianas, unidos entre sí por hileras celulares que le dan continuidad12,13. A pesar de su similitud estructural y microscópica, pueden tener conexiones y características citoquímicas diversas, lo que habla de una multiplicidad de funciones que han sido resumidas en el cuadro precedente. Líneas punteadas: muestran las extensiones más características de estos grupos celulares, que pueden llegar casi hasta la cápsula interna por la lámina medular externa del nervio lenticular o prolongarse a lo largo del lecho de la estría terminal (flecha verde)12,13. Flecha naranja: fascículo prosencefálico medial. El límite interno de la sustancia innominada no es neto y se confunde con las estructuras del túber.

Fig. 14. A. Resumen de las estructuras anatómicas del EPA comparando un corte horizontal tomado de Foix y Nicolesco y nuestro material de disección. Flechas azules: extremo lateral del EPA en la cisura de Silvio, Nc: núcleo caudado, B: banda de Broca. Am: amígdala medial y lateral. Co: cintilla óptica. Ci: cápsula interna. Flecha verde: pedúnculo talámico inferior. Flecha lila: asa lenticular, fo: Campo de Forel, Flechas naranja: fascículo prosencefálico medial. Flechas rojas: pilar del trígono- haz mamilo talámico. n: sustancia negra. r: núcleo rojo. Tcs: tubérculo cuadrigémino superior. La extensión aproximada de la sustancia innominaaa se expresa en el perímetro lila.

Fig. 15. El sistema del pálido ventral, Redibujado y modificado de Haber. El pálido ventral ha sido objeto de investigación en las últimas décadas del siglo pasado11. A pesar de sus límites mal definidos, tiene proyecciones diferentes de las de la parte superior del pálido, como así también aferencias que provienen exclusivamente del estriado ventral, que a su vez, recibe proyecciones exclusivas del sistema límbico. El pálido ventral ocupa una parte pequeña anterior y baja del pálido externo (alrededor de la comisura anterior) y la zona vecina de la sustancia innominada, donde tiene límites poco netos (flechas rojas). El estriado ventral, ocupa la parte baja y anterior del putamen, el puente de sustancia gris que lo une con la cabeza del caudado y parte del nervio accumbens, Hassler lo denominó fundus striati14.
Cuadro 3. Conexiones del pálido ventral

Cuadro 4. Constitución del ganglio basal de Meynert. Cada núcleo parece tener conexiones específicas con un tipo de corteza
 .
.

Fig. 16. Componentes del ganglio basal de Meynert según Arendti16, redibujado y levemente modificado. Se observa la ubicación de los tres grupos chat positivos. Junto al núcleo de la banda diagonal de Broca, hemos agregado en rojo, la ubicación aproximada del pálido ventral según Haber11 aunque los límites entre ambos no son para nada tan netos.
La diferenciación entre núcleo basal y sustancia innominada es, de acuerdo a estos conceptos, solamente histoquímica. Obsérvese la extensión hacia arriba del nervio basal por la lámina medular externa del nervio lenticular. Cada núcleo parecería tener una conexión cortical precisa16. Junto al núcleo de la banda diagonal de broca vemos en rojo la ubicación del pálido ventral aunque los límites entre ambos, no son para nada netos.
Del septum medial y del núcleo de la banda diagonal de Broca sale una proyección que a través del fórnix precomisural y el fórnix dorsal llega al hipocampo siendo el origen de la acetilcolina de este núcleo17,18 ( Alonso y Okeefe, cuadro 5). De la interrelación de estas dos áreas se supone que se genera el ritmo theta actuando como marcapaso probablemente el septum18, Este ritmo invade amplias áreas del lóbulo temporal en algunos individuos.
El núcleo basal de Meynert, alcanza la neocorteza por una proyección difusa unidireccional. El daño de este sistema se ha correlacionado con los trastornos de memoria y otros trastornos cognitivos en la enfermedad de Alzheimer,
La amígdala, por sus conexiones septohipotalámicas y frontobasales, estas últimas a través del núcleo dorsomediano del tálamo, seria un modulador de las respuestas emocionales y de la conducta19. También tendría influencia en los procesos de la memoria episódica relacionados con los componentes emocionales de los eventos20.
Cuadro 5. Conexiones de los componentes del ganglio basal de Meynert

CONCLUSIONES
El EPA es un pequeño lugar cerebral donde se concentran, por un lado, vasos esenciales para el telencéfalo, ya sea "en passage" o penetrantes, y por otro, estructuras nerviosas de importancia funcional relevante para distintas capacidades cognitivas, como la memoria, la motivación, la atención, la personalidad y la conducta.
La disposición de las estructuras nerviosas y de los vasos que las irrigan sigue un plan armónico y topográficamente descriptible, relacionado, posiblemente en parte, con factores filogenéticos.
La comprensión del detalle anatómico y de las relaciones vasculonerviosas, permitirán un manejo quirúrgico de la zona y una interpretación clínica de los cuadros generados por la patología, acordes con la importancia de las estructuras implicadas.
Fe de erratas. Las figuras 13 y 18 de la Parte I de este trabajo han sido tomadas del atlas de De Armond et al21 y no de Foix y Nicolesco como figura en su epígrafe.
Bibliografía
1. Abbie, AA, The morphology of the forebrain arteries, with special reference to the evolution of the basal ganglia, J. Anat, 1934; 68: 433-70.
2. Rosner, SS, Rothon, AL Jr, y col, Microsurgical anatomy of the anterior perforating arteries, J. Neurosurg, 1984; 61: 468-85.
3. Duret, H. Recherches anatomiques sur la circulation de l'encephale. Arch Physio, 1874; 1 : 60- 96.
4. Gillilan, LA. The arterial and venous blood supplies to the forebrain (including the internal capsule) of primates, Neurology, 1968; 18: 653- 70.
5. Salamon, G,, Huang, YP, Radiological anatomy of the brain. Berlin, Springer, 1978.
6. Andersen, PE. The lenticulo-striate arteries and their diagnostic value. A preliminary report. Acta Radiol, 1958, 50: 84- 91.
7. Lazorthes, G, Vascularisation et circulation de léncephale, París, Masson, 1961.
8. Westberg, G, Arteries of the basal ganglia, Acta Radiol. (Diagn.), 1966, 5: 581- 596.
9. Schlesinger, B. The upper brain stem in the human, Springer, Berlin, Heidelberg, NewYork, 1976.
10. Crosby, E. Correlative anatomy of the nervous system, New York The Macmillan Co, 1962.
11. Haber, SN, Lynd-Balta, E,, Mitchell, SJ, The orga nization of the descending ventral pallidal projections of the monkey, J Comp Neurol, 1993; 329: 111-28.
12. Foix,Ch., Nicolesco, J. Les noyaux gris centraux et la région mesencephalo-sous optique, Paris, Mas-son et Cie,, 1925.
13. Jones, EG, Burton, H,, Saper, CB, Swanson, LW, Midbrain, diencephalic and cortical relationships of the basal nucleus of Meynert and associated structures in primates. J Comp Neurol, 1976; 167: 385-420.
14. Hassler,R. Striatal control of locomotion, intentional actions and of integrating and perceptive activity, J Neurol Sciences, 1978; 36: 187-224.
15. Alexander, GE, DeLong, MR, Strick, PL, Parallel organization of functionally segregated circuits linking basal ganglia and cortex, Ann Rey Neurosci, 1986; 9: 357-81.
16. Arendt, T., Bigl, V., Arent, A,, Tennstedt, A, Loss of neurons in the nucleus basalis of Meynert in Alzheimer,s disease, Acta Neuropathol, 1983; 61: 101-8.
17. Alonso, JR, Hoi Sang, U,, Amaral, DG, Cholinergic innervation of the primate hippocampal formation. II, Effects of fimbria/ fornix transection, J Comp Neurol, 1996; 375: 527-51.
18. O'Keefe, J., Nadel, L. The hippocampus as a cognitive map, Oxford, Clarendon Press, 1978
19. Duvernoy, HM, The human hippocampus, Berlin, Heidelberg, Springer, 1998.
20. Markowitsch, HJ. The anatomical bases of memory, En: Gazzaniga, M. The new cognitive Neurosciences. 2a edición. Cambridge (Mass.) MITPress, 2000.
21. De Armond Si, Fusco MM, Dewey, MM , Estructura del Cerebro Humano, Atlas fotográfico, 2a edición, Buenos Aires, Inter- Médica, 1978.